

Professeur d’Histologie à la faculté de Médecine Saint-Antoine, Université Paris VI, en 1968 : cet état fait suite au parcours suivant :
Zoom sur
Exemple de pédagogie: la politique de la vie
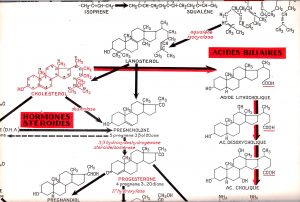
Synopticbiochim
La nécessité où je me trouve de parler à la première personne m’offre du moins l’attrait de rendre hommage à mes Maîtres.
Thèse pour le doctorat en Médecine
«Le rôle de l’utérus dans le contrôle hormonal du relâchement des ligaments pelviens» 20 octobre 1950

coupe d’une corne utérine (x7)

Ligaments pelviens
Directeur adjoint à l’École pratique des Hautes Études près le Collège de France.

Professeur Marois, laboratoire d’histologie, Paris.
En 1945, M. COURRIER, Professeur de « Morphologie expérimentale et Endocrinologie » au Collège de France, me fit l’honneur de m’accueillir dans son laboratoire. Il fut pour moi un Maître dans l’acception la plus noble et la plus riche du terme.
Nommé en 1946 Attaché de Recherches au C.N.R.S. et en 1950, Chargé de Recherches, je quittai la même année le C.N.R.S. Je venais d’être désigné comme titulaire, au poste de Directeur adjoint du Laboratoire de « Morphologie expérimentale et Endocrinologie » de l’École pratique des Hautes Études près le Collège de France.
Les hommes généralités 1 : causerie radiophonique 03/08/1949, Maurice Marois.
Les hommes généralités 2 : causerie radiophonique 10/08/1949, Maurice Marois.
Les hommes généralités 3 : causerie radiophonique 13/08/1949, Maurice Marois
Les hommes généralités 4 : causerie radiophonique 24/08/1949, Maurice Marois.
Les glandes surrénales : causerie radiophonique 14/09/1949, Maurice Marois.
Les glandes surrénales (fin) : causerie radiophonique 21/09/1949, Maurice Marois.
Les hormones : le pancréas. Causerie radiophonique 12/10/1949, Maurice Marois.
I.Un problème d’endocrinologie sexuelle : l’intervention de l’utérus dans le contrôle hormonal du relâchement de la ceinture pelvienne.

Endomètre (x90)
Le relâchement de la ceinture pelvienne marque chez le cobaye la fin de la gestation et rend possible l’accouchement. Le même relâchement s’observe en dehors de la grossesse chez le cobaye ovariectorisé traité par des doses choisies d’œstradiol et de progestérone. L’hystérectomie entraîne la fermeture de la symphyse. Notre démonstration apportait des arguments tirés de la physiologie et de la morphologie expérimentale, en faveur de la spécificité de la relaxine, hormone découverte en 1929, mais dont l’existence fut longtemps contestée.

Myomètre (x90)
Je consacrai à ce problème ma thèse de Médecine ; et l’ensemble de mes recherches sur ce sujet me valut le Prix Montyon de physiologie de l’Académie des Sciences, le Prix Janssen de l’Académie de Médecine, le Prix Châteauvillard et la Médaille d’argent de la Faculté de Médecine de Paris.
Après cette étude de morphologie expérimentale, M. COURRIER m’a proposé d’aborder avec les techniques les plus nouvelles quelques problèmes d’endocrinologie.
II. Radioéléments, endocrinologie et physiologie : fixation de la thyroxine radioactive dans la posthypophyse, éveil de la fonction thyroïdienne, perméabilité capillaire et radiosodium.
C’est ainsi que je fus initié au maniement des radioéléments dans le laboratoire du savant qui avait découvert la radioactivité artificielle. Avec l’équipe des chimistes des laboratoires de Chimie nucléaire et de « Morphologie expérimentale et Endocrinologie » du Collège de France, nous avons étudié le destin physiologique et métabolique de la première hormone marquée à l’aide d’un radioélément, la thyroxine. La découverte de la pénétration élective de la thyroxine dans la posthypophyse couronna cet effort, et je fus désigné pour présenter au Congrès des Endocrinologistes de langue française de 1951 le rapport sur la régulation de la fonction thyroïdienne étudiée à l’aide des radioéléments. Nous avons recherché aussi les relations avec les récepteurs, d’une hormone œstrogène : l’acide dimethyléthylalenolique, marqué avec du radiobrome.
Nous vivions alors la période héroïque de l’application à la biologie de la méthode des radioéléments. A cette époque nous avons étudié avec MM. JOST et MOREL l’éveil de la fonction thyroïdienne de l’embryon de lapin et l’affinité pour le radio-iode de le la thyroïde chez le fœtus décapité. Avec M. MOREL, enfin, nous avons mis au point l’une des premières méthodes d’exploration de la perméabilité capillaire, grâce au radiosodium.
En raison de cette spécialisation précoce, j’ai été appelé à participer très tôt à l’enseignement organisé pour les utilisateurs de radioéléments, par le Commissariat à l’Energie atomique, le Centre national de la Recherche scientifique et l’Institut national d’Hygiène.
III. Morphologie expérimentale, biochimie et endocrinologie.
Après la physique nucléaire, la biochimie est venue collaborer avec la morphologie et l’endocrinologie. M. COURRIER m’a associé à une équipe avec M. Jean ROCHE et son école. La purification de la relaxine, la découverte de la concentration du radio-iode dans le jaune de l’œuf de poule, la démonstration du passage de l’iode radioactif dans le lait, l’étude du métabolisme des protéines iodées, tels furent les premiers fruits de ce travail commun. Ces travaux de recherche pure sur le passage d’un radioélément dans le lait ou sa concentration dans le jaune de l’œuf ont eu des prolongements pratiques importants, car ils ont démontré les dangers de l’entrée dans les cycles biologiques de certains déchets radioactifs.
IV. Sur l’action biologique des rayonnements.
De l’utilisation des radioéléments à l’étude de l’action biologique des rayonnements, il n’y avait qu’un pas : je l’ai franchi en 1953 et je publiai en 1954 avec M. BACLESSE, Chef de Service de la Fondation Curie, ma première communication sur ce sujet. Nous avons démontré que l’hypothermie provoquée par l’administration de chlorpromazine associée au refroidissement augmente la résistance aux rayons X du rat mâle adulte.
J’ai constaté plus tard que ce procédé de l’hypothermie protège l’embryon du rat contre les malformations provoquées par les radiations ionisantes, mais n’est d’aucun secours contre l’apparition tardive de tumeurs.
J’ai présenté au Colloque sur les Défenses de l’Organisme organisé à la Faculté de Médecine de Bordeaux un rapport sur « Les défenses de l’organisme contre les radiations ionisantes ». Le Ministère de la Santé Publique m’a nommé membre de la commission de la protection sanitaire civile et membre correspondant de la commission du Codex (Section des radioéléments).
V. Corticosurrénales et sexualité.
Dès que la chimie offrit à l’expérimentateur la cortisone, M. COURRIER voulut en rechercher l’action éventuelle dans la sphère génitale : nous avons montré son pouvoir sur des récepteurs riches en acide hyaluronique tels la crête du chapon et la peau sexuelle du singe ; nous avons comparé son action sur la corticosurrénale et le tractus génital du singe avec celle de la désoxyxorticostérone. Ces recherches furent décrites dans un rapport de M. COURRIER, de Mme BACLESSE et de moi-même sur « les relations entre la corticosurrénale et la sexualité » présenté devant l’Association des Physiologistes de langue française, en 1953.
VI. Hypothermie et multiplication cellulaire.
Enfin, grâce à une technique originale, mise au point par le physiologiste yougoslave GIAJA, j’ai provoqué l’hypothermie d’un animal homéotherme, le rat ; j’ai recherché le retentissement de l’abaissement de la température centrale de 37° à 18°, sur l’évolution de la grossesse et le développement du fœtus. J’ai constaté qu’un refroidissement quotidien du 1er au 8è jour de la grossesse entraîne un ralentissement du développement de l’embryon qui peut atteindre six jours. Au rétablissement de la température corporelle à 37° le développement des embryons reprend son cours normal. J’ai étendu cette recherche à l’action de l’hypothermie sur la multiplication des cellules tumorales d’un myélome transplantable. J’ai constaté que l’hypothermie ralentit la croissance de la tumeur transplantée et augmente la durée de la survie. Lorsque la tumeur est greffée dans une région froide de l’organisme, l’extrémité de la queue, la tumeur ne se développe plus localement. Si l’extrémité de la queue est insérée dans la cavité abdominale, la croissance de la tumeur reprend.
J’ai enfin découvert le fort pouvoir hypothermisant d’une substance anti-oxydante et réductrice, la vanilline.
VII. Recherches sur quelques problèmes de régulation en endocrinologie.
Nos recherches ont porté sur la régulation des fonctions thyroïdiennes et corticosurrénaliennes.
VIII. Histologie et embryologie.

Laboratoire d’histologie, Paris.
Tels sont quelques-uns des moyens mis en œuvre dans les voies où nous nous sommes engagés. Ces voies et ces moyens peuvent paraître divers. Je voudrais néanmoins tenter de dégager l’unité de l’ensemble.
Certes, les courants de la physique, de la biochimie, de la chimie organique, de la physiologie, ne pouvaient pas laisser indifférente la morphologie expérimentale que nous essayions de servir. Mais les techniques si variées que nous avions utilisées n’avaient, en définitive, qu’un objet : interroger la cellule, le tissu, l’organe, l’organisme. Et, à chaque instant, le recours au microscope nous rappelait que l’histologie est une discipline fondamentale.
Elle m’a révélé que les oestrogènes, en provoquant des nécroses ou des transformations kystiques de l’utérus, avaient opéré une sorte d’hystérectomie hormonale dont la sanction physiologique fut la fermeture de la symphyse pubienne. Elle a montré le parallélisme entre le début de la forte fixation du radio-iode par les thyroïdes embryonnaires et l’apparition du colloïde dans les follicules de la glande. Elle a rendu possibles les études sur la réaction déciduale traumatique et sur les modalités du conditionnement hormonal de ce phénomène. Elle a analysé la réponse du cortex surrénal et du tractus génital de primates soumis à divers traitements hormonaux. Elle a identifié la nature des tumeurs provoquées par les rayons X chez le rat et le passage de certaines formes épithéliales à une forme d’aspect conjonctif. Elle m’a permis d’explorer les réactions de la gonade et du tractus génital de l’embryon du rat dont les mères ont été traitées très précocement par des hormones mâles. C’est à elle que le biochimiste Jean ROCHE nous a demandé de localiser avec précision dans la symphyse pubienne et l’utérus les activités phosphatiques alcalines tandis qu’il en mesurait, par ses propres méthodes, les variations sous l’influence d’hormones sexuelles, chez des animaux normaux ou scorbutiques. Notre analyse simultanée biochimique et histochimique d’activités enzymatiques date de 1949 et 1950, c’est-à-dire du début de l’essor de l’histochimie enzymatique en France.
Ma curiosité pour l’histologie s’est étendue à l’embryologie : étude du conditionnement maternel du développement embryonnaire, exploration de la fonction thyroïdienne fœtale, de l’action sur le fœtus des hormones sexuelles, de l’hypothermie, des rayons X. En 1956, j’ai subi avec succès les épreuves du certificat d’études supérieures d’histologie et de cytologie de la Sorbonne.
La haute récompense de mon mouvement vers l’histologie fut la confiance que m’a témoignée M. le Professeur VERNE. En 1952, M. VERNE me fit l’honneur de m’inviter à faire une conférence sur « l’endocrinologie sexuelle des singes » et en 1955, il me convia à siéger au Conseil de la Société française d’Histochimie qu’il venait de fonder. La publication du rapport sur « l’endocrinologie sexuelle des singes » dans Biologie Médicale m’a valu d’être invité par le Pr. L. LAUNOY, Directeur scientifique de cette revue, à exercer le rôle de Rédacteur en chef, rôle que j’ai continué d’exercer sous la haute direction du Pr. M. GUILLOT. Avec bonheur je constate que MM. COURRIER, ROCHE, VERNE, LAUNOY, GUILLOT étaient tous membres de l’Académie nationale de Médecine.
Professeur agrégé à la Faculté de Médecine.
Le succès au concours d’Agrégation d’Histologie et d’Embryologie des Facultés de Médecine en 1958 m’a fait accéder aux fonctions de Professeur agrégé d’Histologie à la Faculté de Médecine de Paris à partir du 1er octobre 1958 puis à la Faculté de Médecine Saint-Antoine de l’Université Paris VI, dès l’ouverture de ce premier C.H.U. sous le décanat de M. le Professeur André LEMAIRE. En même temps que j’assumai les charges hospitalo-universitaires, je poursuivais mes recherches histophysiologiques en endocrinologie sexuelle.
I. Étude de divers stéroïdes progestatifs de synthèse.
Les progestatifs ont pour premier pouvoir celui de maintenir la grossesse. Mais les progestatifs de synthèse peuvent présenter certaines propriétés secondaires androgènes et oestrogènes qui font courir le risque d’une masculinisation ou d’une féminisation des fœtus. Nous avons recherché les propriétés secondaires des principaux stéroïdes progestatifs, dont certains sont anticonceptionnels (norethindrone, chlormadinone, medroxyprogestérone, mégestrol, diméthistérone, norethynodrel, allylestrenol, dydrogestérone) et mis au point sur le fœtus de rat des tests histométriques d’une grande sensibilité. Ces tests permettent d’établir si un stéroïde est doué d’un pouvoir masculinisant ou féminisant ou des deux pouvoirs simultanément.
II. Étude de l’action progestative indirecte de certains inhibiteurs de la stéroïdogenèse corticosurrénalienne.
Chez la ratte ovariectomisée traitée par l’amphénone, nous avons observé que la corticosurrénale acquiert le pouvoir de maintenir la gestation. Sous l’empire de l’amphénone, la glande a donc sécrété des substances progestatives. Ainsi avons-nous pu confier pour la première fois à une glande jouant un rôle important dans la physiologie de l’individu une fonction de l’espèce : le maintien de la grossesse. Chemin faisant, nous avons réalisé l’étude histochimique de la corticosurrénale de la ratte gravide traitée par l’amphénone, et de ses fœtus. Et nous avons recherché l’action éventuellement progestative d’un autre inhibiteur de la stéroïdogenèse, le SU 4885.
III. Les hormones sexuelles maternelles retentissent-elles sur la différenciation sexuelle du fœtus ?
Nous avons reproduit la délicate technique de la castration fœtale chez le lapin et constaté que les sécrétions ovariennes de la mère n’étaient pas responsables du maintien du canal de MULLER chez l’embryon.
IV. Androgènes, gestation et fœtus.
Nous avons découvert que dans certaines circonstances, l’ovaire peut exercer une action novice sur la gestation et sur les fœtus. Injecté à des rates gravides entre le 7è et le 11è jour, un androgène, le dipropionate d’androsténediol, provoque l’avortement ; injecté en fin de gestation, il détermine une forte réduction de la croissance fœtale. Ces actions ne se manifestent plus chez des rattes ovariectomisées dont la gestation est maintenue par la progestérone ; ainsi, en l’absence d’ovaires, l’effet néfaste disparaît. Nous avons étendu ces observations à de nombreux androgènes et constaté la particulière nocivité des androgènes dont le cycle A est susceptible d’être aromatisé par l’ovaire : l’androgène est alors aromatisé en œstrogène. Cette capacité de l’ovaire de transformer un androgène en œstrogène avait été démontré chez la femme dans une publication princeps de Guy LAROCHE, H. SIMONNET et E. BOMPARD (1939).
Afin de démontrer par une autre voie le rôle de l’ovaire, nous avons réalisé une sorte de « castration chimique » par un détergent, le triton : il provoque un déficit de la fonction lutéale. La testostérone perd alors son pouvoir abortif comme chez l’animal ayant subi une ovariectomie chirurgicale.
V. Sur le déterminisme de l’accouchement.
Les facteurs qui provoquent le déclenchement de la parturition n’étaient pas entièrement connus. La sérotonine avait été mise en cause. Nos observations sur la ratte gestante nous autorisent à dénier à la sérotonine un rôle important. Chemin faisant, nous avons mis au point une méthode expérimentale permettant de tester l’action in vivo de la sérotonine et de ses antagonistes. Ce test est désormais utilisé dans de nombreux laboratoires.
Professeur à la Faculté de Médecine.
En 1968, j’eus l’honneur d’accéder à la fonction de Professeur d’Histologie à la Faculté de Médecine Saint-Antoine de l’Université Paris VI. J’ai poursuivi mes activités enseignantes et hospitalières en même temps que je développais mes travaux de recherche.
I. Recherches sur les œstrogènes faibles et les antiandrogènes.
A) Œstrogènes faibles.

Recherche
Un groupe de substances, les œstrogènes faibles, joue un rôle important comme instrument de recherche en endocrinologie expérimentale et comme agent thérapeutique. Deux catégories d’entre elles ont suscité un intérêt particulier : les œstrogènes entravés du type de l’oestriol, par exemple le fridéron, et les anti-œstrogènes du type du clomifène ou du tamoxifène agissant par compétition avec les oestrogènes sur les récepteurs.
-
- Travaux avec un œstrogène entravé : le fridéron.Le fridéron est un œstrogène entravé qui fut utilisé contre les troubles de la ménopause à cause de l’inhibition qu’il exerce sur la sécrétion de FSH. Nous avons démontré cette inhibition sur le test de la fonte testiculaire du rat par atteinte de la spermatogenèse ; en contre épreuve, nous avons empêché cette fonte grâce à l’injection de PMS (sérum de jument gravide riche en FSH). Nous avons confirmé les faibles activités œstrogènes du fridéron sur le vagin, l’utérus et la mamelle du rat. Nous avons enfin observé que l’effet utérotrophe de cette substance comme de tout œstrogène entravé présente une courbe dose réponse de pente plus faible que l’œstradiol.
- Travaux avec des anti-œstrogènes.
a) Le clomifène : Le clomifène est un analogue chimique d’un œstrogène de synthèse, le chlorotrianisène.
-
-
-
- Action sur les follicules ovariens : nous avons découvert que le clomifène augmente chez la lapine le nombre de follicules primaires polyovulaires. Cette expérience soutient l’hypothèse de relations inverses entre le nombre de follicules polyovulaires et le taux d’œstrogène circulant.
- Action sur divers récepteurs sexuels et dans diverses circonstances physiologiques : nous avons étudié l’action du clomifène sur le vagin et l’utérus de la ratte et sur la symphyse pubienne du cobaye. Nous avons aussi étudié ses effets sur le déciduone traumatique, la nidation, l’évolution de la grossesse et l’accouchement chez la ratte, et l’action antagoniste de l’aromatisation de certains androgènes en prenant pour tests dans la même espèce le retentissement sur la gestation et sur la différenciation sexuelle somatique.
-
-
b) Le tamoxifène : Le tamoxifène est un dérivé d’un œstrogène de synthèse non stéroïde, le triphényl éthylène. Il a été utilisé en thérapeutique humaine contre les carcinomes mammaires oestrogéno-dépendants. Nous avons observé chez la ratte que cette substance est œstrogène faible et antioestrogène (antagoniste de fortes doses d’œstradiol) sur le vagin, l’utérus et la glande mammaire (développement, ouverture et sécrétion des acini).
B) Antiandrogènes.
Les antiandrogènes du type de la cyprotérone offrent un précieux instrument d’analyse du déterminisme de la différenciation sexuelle. Nos résultats chez le fœtus mâle du rat montrent qu’administrée à la mère au cours de la gestation, la cyprotérone empêche la masculinisation du tubercule génital et de la distance ano-génitale et fait apparaître un bulbe sino-vaginal, mais laisse intacts les canaux de WOLFF ; elle est impuissante à s’opposer à la régression des canaux de MULLER. La cyprotérone ne s’oppose pas à l’action nocive de propionate de testostérone sur la gestation du rat mais elle est antagoniste de l’action masculinisante de cette hormone sur certains territoires du tractus génital du fœtus femelle.
II. Retentissement de l’administration des ions cuivres et zinc sur les fonctions ovariennes.
Sur l’ovulation de la lapine, nous avons confirmé le pouvoir inducteur de l’ion cuivre, l’inefficacité de l’ion zinc injecté seul et nous avons démontré une synergie des deux ions. Le zinc exercerait une action périphérique en augmentant la durée d’efficacité des hormones gonadotropes circulantes alors que le cuivre exerce ses effets sur les centres de contrôle hypothalamique des fonctions gonadotropes.
Chez la ratte, nous avons constaté que l’acétate de cuivre permet d’interroger la capacité réactionnelle du système hypothalamo-hypophysaire dans ses relations avec l’ovaire. Dans les races de rats chez lesquels un éclairement continu provoque facilement un rut permanent, une injection d’acétate de cuivre ne transforme pas le corps jaune cyclique en corps jaune de pseudo-grossesse. Cette transformation s’opère lorsque l’administration du cuivre est répétée plusieurs jours ; nous avons observé dans cette circonstance une synergie avec l’ion zinc.
Enfin, à la différence de la gestation de la lapine, la gestation de la ratte est interrompue par l’administration quotidienne d’acétate de cuivre. La progestérone exerce une action protectrice.
III. Drogues psychotropes, fonctions ovariennes et activité mammaire.
1. De nombreuses observations cliniques et expérimentales ont montré que certains dérivés de la phénothiazine provoquent une galactorrhée. Cet effet est attribué à la libération de la prolactine, ces dérivés de la phénothiazine agissant sur le contrôle hypothalamique de la sécrétion de cette hormone.
Nous avons comparé chez la ratte adulte l’action de trois dérivés des phénothiazines (thiopropérazine, trifluopérazine, chlorpromazine) à celle de la réserpine. Ces quatre drogues psychotropes ont pour des doses croissantes des effets croissants, le nouvel effet se surajoutant au précédent : installation d’un di-oestrus puis formation d’un déciduome, enfin sécrétion mammaire. La thiopropérazine s’est montrée particulièrement active.
2. Nous avons retenu cette substance pour les expériences ultérieures. La thiopropérazine injectée chez la ratte au début de la gestation retarde la nidation ; administrée du 7è au 10è jour, elle entraîne la résorption des fœtus. Ces deux effets sont corrigés par l’administration de faibles doses d’œstradiol. La thiopropérazine a donc inhibé la sécrétion de FSH.
3. Des œufs prélevés le jour 5 de la gestation dans le corne d’une ratte gravide et transférés dans des cornes de femelles rendues pseudo-gestantes par la thiopropérazine s’implantent et se développent jusqu’à terme ; l’accouchement et l’allaitement sont normaux.
4. Un ovaire de ratte greffé dans le ligament large peut continuer d’exercer ses fonctions : les rattes présentent un cycle régulier 15 à 25 jours après la greffe. Rendues pseudo-gestantes, soit par coït, soit par administration de thiopropérazine, elles tolèrent l’implantation d’œufs transplantés le 5è jour dans leurs cornes ; la gestation se poursuit jusqu’à son terme : l’accouchement et l’allaitement sont normaux comme dans les expériences précédentes.
IV. Prostaglandines et déterminisme de l’accouchement.
1. Les prostaglandines ont été impliquées dans les mécanismes de l’accouchement. Avec les découvreurs de ces substances : VON EULER, BERGSTROM et SAMUELSON de Suède, j’ai organisé la première conférence mondiale sur ces composés nouveaux.
2. La diphényle paraphénylénediamine (DPPD) est un anti-oxydant. Nous avons démontré que le retard de l’accouchement qu’elle provoque chez la ratte est renversé par la prostaglandine F2alpha. La DPPD a sans doute inhibé la synthèse des prostaglandines. L’enseignement de cette recherche est l’éventuel intérêt des anti-oxydants contre la prématurité.
V. Vieillissement et sécrétion lactée.
Chez la ratte, un éclairement permanent ou la répétition de stress ne retentit pas sur la mamelle de femelles jeunes mais provoque toujours chez la femelle âgée de dix-huit mois une abondante lactation. Ainsi, le vieillissement associé aux modifications de l’environnement entraîneraient une sécrétion augmentée de prolactine.
VI. Sur l’administration d’une tétracycline pendant la gestation ou dans les semaines qui suivent la naissance : fixation dans le squelette ou la dent par chélation du calcium.
Les tétracyclines chélatent les ions métalliques bivalents en particulier le calcium et forment ainsi des complexes plus ou moins stables. Cette propriété explique leur affinité pour les tissus calcifiés : leur persistance est en corrélation directe avec le contenu en calcium des territoires considérés.
Nous avons recherché l’action de la demethyl tétracycline (DMTT) sur la croissance des incisives et du fémur du rat, le produit étant administré à la mère gestante ou allaitante ou aux jeunes rats de quatre jours ou d’un mois. Pour la mise en évidence de la fixation de la DMTT, nous avons pratiqué l’examen au microscope de la fluorescence en lumière ultraviolette. La fluorescence des tissus minéralisés du squelette et de la dent est due au complexe tétracyclines-calcium. Il est important de comprendre le mécanisme du dépôt de la tétracycline dans l’os pour éclairer les relations entre les substances minérales et la matrice protéique. Une telle recherche aidera peut-être à mieux connaître le processus de fixation dans le squelette de certaines substances ostéotropes radioactives tel le radio strontium dont la présence dans l’os n’est pas souhaitable. Je dois à ces travaux le titre de membre de l’Académie nationale de Chirurgie Dentaire.
VII. Sur quelques glandes sécrétrices de phéromones.
Les phéromones sont des signaux olfactifs qui jouent un rôle important dans les comportements sexuels et sociaux. La sécrétion de certaines d’entre elles est sous l’empire des hormones sexuelles.
Chez les animaux normaux et chez des animaux traités ou non par des hormones sexuelles mâles ou femelles, nous avons réalisé une étude histochimique des glandes anales et inguinales du lapin et des glandes bulbouréthrales et préputiales du rat.
Nous avons accordé une attention particulière aux glandes préputiales du rat à cause de leur intérêt morphologique (glandes phéromones). Nous avons observé un dimorphisme sexuel chez le fœtus et décrit la sensibilité des glandes fœtales aux hormones sexuelles mâles ou femelles injectées à la mère. Après la naissance, nous avons constaté que le développement de ces glandes, comparable dans les deux sexes, atteint son maximum dès avant la puberté et régresse chez l’animal vieillissant. Après la castration le poids de ces glandes diminue quel que soit le sexe. Leur réponse aux hormones sexuelles est voisine chez les animaux prépubères ou adultes, entiers ou castrés, mâles ou femelles ; les hormones se classent par activité décroissante dans l’ordre suivant : testostérone, progestérone ; la réponse à l’œstradiol met en jeu l’antéhypophyse.
Les glandes préputiales de souris offrent un intérêt particulier car le sébum qu’elles sécrètent est de même nature que le sébum humain. Nous avons étudié leur développement et leur réaction à divers androgènes (propionate de testostérone, adrosténedione, androstérone) ; nous avons découvert l’action antagoniste contre certains androgènes exercée par des sels de zinc ou un détergent, la supérinone. Ces travaux ouvrent de nombreuses perspectives ; ils peuvent offrir un intérêt pour la recherche de substances susceptibles de s’opposer à la chute des cheveux puisque dans l’appareil pilo-sébacé il existe un équilibre entre glande sébacée et pousse du poil.
VIII. Sur l’intérêt de l’étude de l’embryotoxicité des substances chimiques.
ROBSON a souligné l’importance pour la chimiothérapie des cancers, du test de l’embryotoxicité : toute substance embryotoxique directe mérite d’être éprouvée aussi sur des tumeurs transplantées.
Nous avons démontré l’action toxique sur les fœtus du rat par effet direct d’un colorant des graisses, le soudan II (C18 H16 N2 0 = xylène azo beta naphtol), administré à la ratte gestante du 7è au 10è jour de la grossesse. Nous avons constaté aussi que cette même substance exerce une action inhibitrice de la croissance d’une tumeur transplantée chez le rat : le myélome T58 de GUERIN. Il est possible que le soudan II interfère avec le métabolisme des lipides sur lesquels il se fixe ; l’un des points d’impact pourrait être les lipides des membranes.
Conclusion.
Cet exposé n’aurait dû être qu’un bref curriculum vitae. Mais les étapes de ce curriculum s’identifient à l’histoire de ma recherche et je n’ai pas pu les dissocier. J’ai eu le souci d’élargir ma base de départ en poursuivant mes études parallèlement dans les Facultés de Médecine et de Sciences. Puis, je fus favorisé du destin par la qualité des Maîtres qui m’ont accepté comme élève. Je m’essaye, dans les limites de ce temps d’une vie, à déchiffrer quelques aspects de l’immense problème de la Vie. Connaître le dernier mot, selon le rêve du philosophe ou du mystique, n’est pas à ma portée. Dans cette approche difficile d’une réalité infiniment riche et complexe, je bénéficie de l’effort collectif de la science. J’y participe avec passion. Je ne retire de mon labeur qu’une vision partielle ; elle suffit à me donner une joie profonde qui sous-tend mon action. Cette action ne se donne pas seulement pour objet l’acquisition de connaissances nouvelles en biologie et en médecine mais la diffusion des connaissances acquises : tel fut le sens de mon engagement au service de la Faculté de Médecine.
Mais je ne me sentirais pas quitte envers moi-même si, ayant accompli mon devoir d’état, je ne tentais pas d’assumer aussi des responsabilités sociales. J’assume ces responsabilités au soin d’institutions qui honorent certaines valeurs de civilisation :
– l’Association des Lauréats du Concours général (Secrétaire général en 1966, puis Vice-Président chargé du Secrétariat général depuis 1973).
– la Société française de Thanatologie (Président depuis 1970 puis Président international jusqu’en 1990).
– l’Institut de la Vie créé en 1960 (Président Fondateur, puis au niveau international : Délégué Général)
Liste des publications scientifiques (1947-1998)
magot mâle impubère folliculinisé;réaction épithéliale
1. R. COURRIER et M. MAROIS
La relaxine existe-t-elle ?![]()
C. R. Soc. Biol., 1947, 141, 1202-1203
2. F. MOREL et M. MAROIS
La perméabilité capillaire au sodium, mesurée chez le lapin à l’aide du radiosodium.![]()
C. R. Soc. Biol., 1948, 142, 1366-1369
3. M. MAROIS
Action locale de la progestérone sur la corne utérine et relâchement de la symphyse pubienne du cobaye.![]()
C. R. Soc. Biol., 1948, 142, 1407-1409
4. F. MOREL et M. MAROIS
Méthode de mesure du taux des échanges de sodium entre le plasma et les liquides extravasculaires chez le lapin à l’aide du radiosodium.![]()
Arch. des Sciences Physiol., 1949, 3, 15-26
5. A. JOST, F. MOREL et M. MAROIS
Données préliminaires sur la fixation de radio-iode i131 par la thyroïde foetale du lapin.![]()
C. R. Soc. Biol., 1949, 143, 142-145
6. M. MAROIS
Déciduome traumatique, symphyse pubienne et rapport oestradiol- progestérone chez le cobaye.![]()
C. R. Soc. Biol., 1949, 143, 370-372
7. F. MOREL et M. MAROIS
Actions de l’histamine sur le système vasculaire du lapin, étudiées au moyen de radiosodium.![]()
C. R. Soc. Biol., 1949, 143, 464-468
8. M. MAROIS
Utérus et relâchement de la symphyse pubienne du cobaye.![]()
Bull. Ass. Anat., 1949, 58, 467-474
9. R. COURRIER, J. ROCHE, G. H. DELTOUR, M. MAROIS, R. MICHEL et F. MOREL
Sur la préparation et le métabolisme d’iodocaséines radioactives .![]()
Bull. Soc. Chim. Biol., 1949, 31, 1029-1034
10. J. ROCHE, R. COURRIER, G. H. DELTOUR, M. MAROIS, R. MICHEL et F. MOREL
Sur la préparation et le métabolisme d’iodocaséines radioactives d’inégale activité biologique.![]()
11. R. COURRIER, J. ROCHE, G. H. DELTOUR, M. MARDIS, R. MICHEL et F. MOREL
Sur l’excrétion mammaire d’iode radioactif après administration d’iodures ou d’iodocaséine marqués.![]()
C. R. Soc. Biol., 1949, 143, 599-601
12. R. COURRIER, A. HOREAU, M. MAROIS et F. MOREL
Etude quantitative de la pénétration de la radiothyroxine dans les cellules hypophysaires.![]()
C. R. Soc. Biol., 1949, 143, 935-937
13. R. COURRIER, A. HOREAU, J. JACQUES, M. MAROIS, F. MOREL et P. SUE
Radioendocrinologie, hormones radioactives et récepteurs.![]()
C. R. Acad. Sciences, 1949, 229, 275-277
14. B. NATAF et M. MAROIS
Activité phosphatasique alcaline et peptidasique et ouverture expérimentale de la symphyse pubienne du cobaye.![]()
C. R. Soc. Biol., 1949, 143, 1588-1590
15. M. MAROIS
Le rôle de l’utérus dans le contrôle hormonal du relâchement des ligaments pelviens.![]()
Thèse de Doctorat en Médecine, Paris, 1950, 175 pages
16. M. MAROIS, B. NATAF et P. MAROIS
Méthode radiographique de mesure du relâchement de la symphyse pubienne et modalités de son déclenchement hormonal.![]()
Ann. Endocrinol., 1950, 11, 482-490
17. J. ROCHE, B. NATAF, M. MAROIS
Actions de l’oestradiol et de la progestérone sur les activités phosphatasique alcaline et peptidasique du ligament pelvien et évolution de la symphyse pubienne chez le cobaye.![]()
Ann. Endocrinol., 1950, 11, 491-503
18. B. NATAF et M. MAROIS
Scorbut et relâchement expérimental de la symphyse pubienne du cobaye.![]()
C. R. Soc. Biol., 1950, 144, 1627-1629
19. R. COURRIER et M. MAROIS
A propos des problèmes de l’hormonologie sexuelle.![]()
Schweiz. Med. Wschr., 1951, 81, 989
20. R. COURRIER, A. HOREAU, M. MAROIS et F. MOREL
Sur la pénétration de la thyroxine dans le lobe postérieur de
l’hypophyse. ![]()
C. R. Acad. Sciences, 1951, 232, 776-778
21. J. ROCHE, B. NATAF et M. MAROIS
Scorbut et déclenchement hormonal du relâchement de la symphyse pubienne chez le cobaye.![]()
Ann. Endocrinol., 1951, 12, 212-227
22. M. MAROIS
Quelques aspects de la régulation de la fonction thyroïdienne étudiés à l’aide de radioéléments. (Rapport présenté au 1er Congrès des Endocrinologistes de langue française 1951).![]()
Ann. Endocrinol., 1951, 12, 333-359.
23. M. MAROIS et B. NATAF
Variations simultanées des activités phosphatasique alcaline et peptidasique de l’utérus et ouverture expérimentale de la symphyse pubienne chez le cobaye.![]()
C. R. Soc. Biol., 1951, 145, 1758-1760
24. J. ROCHE, O. MICHEL, R. MICHEL et M. MAROIS
Sur les modalités de la concentration du radio-iode (Iode 131) dans le vitellus de l’oeuf de poule et sur la formation ovarienne de combinaisons iodées.![]()
C. R. Soc. Biol., 1951, 145, 1833-1836
25. M. MAROIS et F. MOREL
Thyroxine marquée, récepteurs et interrelations endocriniennes .![]()
Arch. Anat. Histol . et Embryol . , 1951-1952, 34, 275-284
26. R. COURRIER et M. MAROIS
Relations entre la testostérone et la cortisone dans leurs effets sur certains organes récepteurs .![]()
C. R. Acad. Sciences, 1952, 234, 271-273
27. M. MAROIS et Ch. COMBESCOT
Sur les rapports de la cortisone et de la progestérone envisagés chez la lapine.![]()
C. R. Acad. Sciences, 1952, 146, 838-840
28. Ch. COMBESCOT et M. MAROIS
Sur les rapports de la cortisone et de la progestérone envisagés chez la lapine.![]()
C. R. Acad. Sciences, 1952, 234, 2484-2485
29. M. MAROIS
L’endocrinologie sexuelle des singes.![]()
Biologie Médicale, 1952, 41, 465-506
30. A. JOST, F. MOREL et M. MAROIS
Nouvelles recherches à l’aide du radio-iode 131 sur la fonction thyroïdienne du foetus de lapin décapité.![]()
C. R. Soc. Biol., 1952, 146, 1066-1070
31. M. MAROIS
Contrôle hormonal de la ceinture pelvienne et endocrinologie sexuelle. Le problème de la relaxine.![]()
Masson Edit. Paris, 1953, 175 pages, 16 planches.

